Project in collaboration with the Kondrashov Lab
Over 10 millions invertebrate species are thought to be living on earth. The behaviors of each of them are remarkably complex, mostly innate and controlled by neuronal networks with a unique set of embedded algorithmic rules. These behaviors have evolved for over 500 Myr, an estimated time point of their first speciation event (A). Remarkably, families that diverged around 400 Myr ago, such as Diptera, Hymenoptera and Lepidoptera share a remarkable similarity in their neuronal circuits. In particular, the region of the brain involved in the detection and processing of visual motion – their optic lobes. Two neuropils in the insect’s optic lobes, the lamina and the medulla, appear to be morphologically highly conserved (B-D). The degree of correspondence allows to distinguish all neuronal cells types composing these structures unambiguously between species. Although the physiological properties across species have not been characterized so far, their morphological similarity is a good indication that their functional properties are largely unchanged. In line with this idea, responses to visual stimuli in cells known to integrate the computations of medulla circuitry, the so-called lobula plate tangential cells (LPTCs), show changes in spatio-temporal tuning, but not in their characteristic motion-sensitive responses across species that diverged over 400 Myr ago (E)). The LPTCs, called by some the “cockpit of the fly”, form a network of ~ 30 cells per hemisphere located in the posterior part of the lobula complex, with every single cell-type defined by a distinct morphology and connectivity within that network (C, G-H). Although the presynaptic elements to LPTC are morphologically indistinguishable, LPTCs show drastic morphological changes, even when only comparing across Diptera (E). In collaboration with Fyodor Kondrashov and his lab we are establishing a framework to uncover the role of these evolutionary adaptations and shed light on circuit evolution, development, computations, and behavioral specializations.
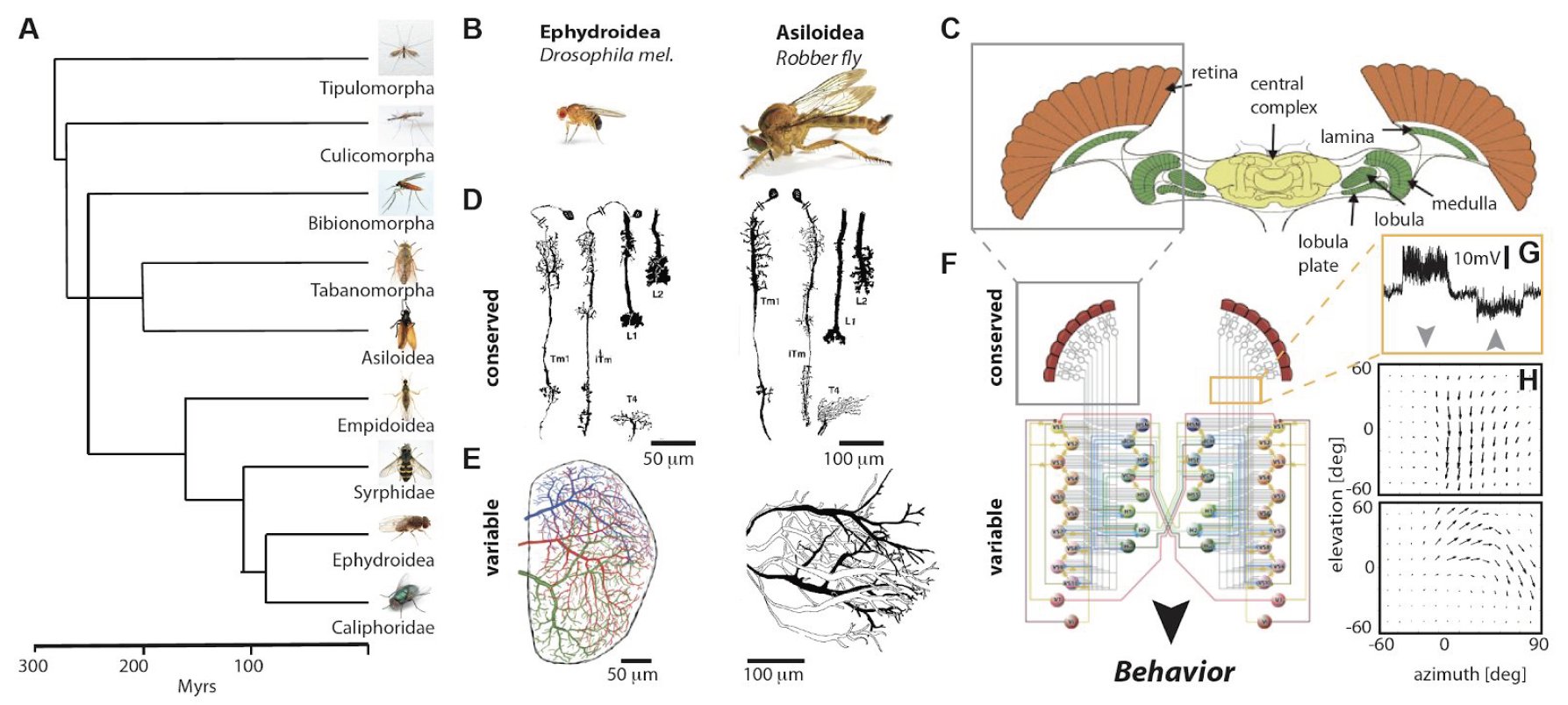
Evolution of visual processing across Diptera. A. Phylogenetic tree of Diptera. B. Two example suborder species exemplifying their conserved and variable neuronal structures in their optic lobes. C. Schematic of the fly brain. D. Morphology of conserved laminar and medullar interneurons across species. E. Morphology of lobula plate tangential cells (LPTCs) across species exemplifying the variability of these neuronal structures. F. Schematic of the course control circuitry in flies. The conserved local motion detection circuitry (grey box) enables the detection of moving objects in a direction-selective manner, as exemplified by the response to a moving grating processed by these motion detectors and integrated by an LPTC (orange box and G, arrow depict the direction of the moving stimulus). These local motion detectors are processed by a network of LPTCs (bottom circuit) that enables LPTCs to have cell-specific network properties, e.g. their optic flow fields (H) to ultimately instruct visually guided behaviors. ((D) modified from (1), (E) from (2,3), (C) from (4) and (F) from (5))
1. Buschbeck, E. K. & Strausfeld, N. J. Visual motion-detection circuits in flies: small-field retinotopic elements responding to motion are evolutionarily conserved across taxa. J. Neurosci. 16, 4563–4578 (1996).
2. Buschbeck, E. K. & Strausfeld, N. J. The relevance of neural architecture to visual performance: phylogenetic conservation and variation in Dipteran visual systems. J. Comp. Neurol. 383, 282–304 (1997).
3. Schnell, B. et al. Processing of horizontal optic flow in three visual interneurons of the Drosophila brain. J. Neurophysiol. 103, 1646–1657 (2010).
4. Borst, A. & Haag, J. Neural networks in the cockpit of the fly. Journal of Comparative Physiology A 188, 419–437 (2002).
5. Borst, A., Haag, J. & Reiff, D. F. Fly motion vision. Annu. Rev. Neurosci. 33, 49–70 (2010).